Un ancêtre potentiel de la Perche nilotique, Lates Niloticus (L)
Identifié à l'état fossile aux environs du Boulou (Pyrénées-Orientales)
Par Jean Gaudant, Paris (UMR 8569 CNRS)
Résumé
Le réexamen de deux poissons fossiles provenant des «grès de Moulas» a montré qu'ils appartiennent au genre actuel Lates Cuvier & Valenciennes, dont les plus anciens représentants actuellement connus sont datés du Miocène. Cela conduit donc à rajeunir considérablement la formation des «grès de Moulas».
Abstract
The reexamination of two fossil fishes from the «grès de Moulas» has shown that they really belong to the recent genus Lates Cuvier & Valenciennes, the oldest representatives of which are known from the Miocene. This leads one to a considerable rejuvenation of the «grès de Moulas» Formation.
Introduction
Au début du siècle dernier, Depéret (1912) tenta d'utiliser quelques squelettes de poissons fossiles pour dater les grès à pavés qui étaient exploités en carrières à Moulas, près du Boulou (Pyrénées-Orientales). Il en confia l'étude à Sauvage qui crut y reconnaître, sans doute en raison de leur grande taille, l'espèce Cyclopoma gigas Agassiz de l'Eocène moyen du célèbre gisement du Monte Bolca, près de Vérone (Italie). Pour cette raison, les «grès de Moulas» furent donc attribués à l'Eocène moyen.
Deux spécimens incomplets provenant des anciennes carrières de Moulas sont conservés au Muséum d'histoire naturelle de Perpignan. On peut raisonnablement penser qu'ils faisaient partie du matériel examiné par Depéret et Sauvage. A l'occasion de la préparation de la feuille de Céret de la carte géologique de France à 1/50000, il a paru utile de les réétudier. Leur récent réexamen (Gaudant, 1999) a permis d'en donner la description suivante.
Description anatomique
 |
| Fig. 1. - Squelette incomplet de Lates provenant des «grès de Moulas», Le Boulou (Pyrénées-Orientales). |
Le premier spécimen (fig. 1) est dépourvu de région caudale. Sa distance antéanale égale approximativement 265 mm, ce qui conduit à estimer sa longueur standard à au moins 370 mm. Le corps est allongé, sa hauteur maximale étant comprise environ 3,5 fois dans la longueur standard estimée. La tête est grande : sa longueur, qui excède la hauteur maximale du corps, est comprise environ 3,4 fois dans cette même dimension. Elle est relativement allongée : sa hauteur maximale égale approximativement les 3/4 de sa longueur. Son état de conservation médiocre ne permet pas d'en donner une description détaillée. On notera toutefois que le toit crânien est surmonté par une crête supraoccipitale longue et basse qui prend naissance au-dessus du milieu de l'orbite et dont la longueur égale 64 % de celle du neurocrâne.
La cavité buccale est modérément développée, la longueur de la mandibule égalant environ la moitié de celle de la tête. De ce fait, son articulation avec le crâne prend place approximativement au-dessous du milieu de l'orbite. Le bord oral du dentaire est garni de nombreuses petites dents villiformes. La mâchoire supérieure comporte un prémaxillaire dont le processus oral, relativement court, ne constitue pas la totalité du bord oral de la mâchoire. Le maxillaire est relativement massif puisque la largeur de son extrémité postérieure égale environ le quart de sa longueur. Il est surmonté par un fort supramaxillaire.
La partie postérieure de la tête est très médiocrement conservée. Pratiquement rien ne subsiste du préopercule. En revanche, l'opercule présente une forme générale subtriangulaire. Son bord postérieur est orné d'une épine unique. Sa largeur maximale paraît avoir égalé environ les 2/3 de sa plus grande hauteur.
Le corps. - La colonne vertébrale paraît être composée de 11 vertèbres abdominales possédant des centra robustes. Les plus antérieurs, qui semblent avoir été relativement courts, sont médiocrement conservés. Les centra suivants sont tous sensiblement allongés, leur longueur égalant approximativement une fois et demie leur hauteur.
Les centra vertébraux abdominaux supportent des neurapophyses dont les deux plus antérieures sont très redressées alors que les suivantes s'inclinent progressivement pour former un angle d'environ 50° avec l'axe de la colonne vertébrale.
Les côtes pleurales sont relativement mal conservées. Il est cependant possible d'en dénombrer au moins 7 paires. Les côtes pleurales postérieures s'articulent avec de longues parapophyses.
Seuls douze des quatorze centra vertébraux postabdominaux peuvent être observés, le fossile étant brisé peu en avant de l'extrémité postérieure de la colonne vertébrale.
La nageoire dorsale est insérée un peu en arrière de la tête. Dédoublée, elle débute par deux courtes épines relativement fines en arrière desquelles prend place une longue épine robuste qui est la plus longue de la nageoire. Sa longueur est un peu inférieure aux 3/4 de la hauteur maximale du corps et égale 1/5 de la longueur standard. Plus en arrière subsistent les restes incomplets de trois épines. En revanche, rien ne subsiste de la dernière épine de la nageoire dorsale antérieure dont l'axonoste proximal correspondant est cependant présent. La nageoire dorsale postérieure débute par une épine assez courte qui est également relativement grêle. Postérieurement prennent place onze lépidotriches dont seul le plus antérieur n'est pas bifurqué.
L'endosquelette de la nageoire dorsale n'est pas intégralement conservé. On constate cependant que 7 ptérygophores sont disposés au-dessous de la dorsale antérieure. En revanche, il n'a pas été possible de déterminer combien de ptérygophores prenaient place au-dessous de la dorsale postérieure.
La nageoire anale est opposée à la moitié postérieure de la dorsale postérieure. Les épines situées à sa partie antérieure n'ont laissé aucune trace. On dénombre en revanche 8 lépidotriches, tous à la fois articulés et bifurqués. Le nombre total de rayons de l'anale devait donc être de 9. L'endosquelette de l'anale n'est pas fossilisé.
De la ceinture scapulaire ne subsiste qu'une partie du cleithrum dont l'angle postéro-ventral n'est pas observable. La nageoire pectorale est réduite aux débris de quelques rayons dont seul l'article basal est conservé.
Seuls des restes fragmentaires des os pelviens sont visibles. De même, quelques débris de rayons témoignent de l'existence des nageoires pelviennes.
 |
| Fig. 2. - Squelette incomplet de Lates provenant des «grès de Moulas», Le Boulou (Pyrénées-Orientales). |
Le second spécimen (fig. 2) est dépourvu de tête mais permet en revanche d'observer la région caudale dans son ensemble. Il s'agit d'un individu sensiblement plus grand que le précédent car sa longueur standard devait être voisine de 465 mm si l'on compare la longueur de sa région postabdominale (mesurée depuis l'extrémité antérieure du premier centrum postabdominal jusqu'au bord postérieur des hypuraux) qui égale 205 mm, à celle observée chez l'espèce Lates aquensis Gaudant, de l'Oligocène terminal d'Aix-en-Provence.
La colonne vertébrale est incomplète puisque seuls huit centra abdominaux sont observables. On constate en revanche que la région postabdominale est constituée de 14 vertèbres. Les trois dernières contribuent à soutenir la nageoire caudale.
Les côtes pleurales, au nombre de 9 paires, sont relativement courtes puisque leur extrémité distale se situe approximativement au tiers inférieur de la cavité abdominale, mesurée de la région inférieure des centra au bord ventral du corps. Les deux paires postérieures de côtes sont portées par des parapophyses relativement longues.
Bien que l'extrémité distale de ses rayons ne soit pas conservée, on peut affirmer que la nageoire caudale avait une forme en palette. Elle est constituée de 17 rayons principaux dont 15 sont à la fois articulés et bifurqués. En avant de ces rayons prennent place, dorsalement et ventralement, 7 ou 8 rayons marginaux.
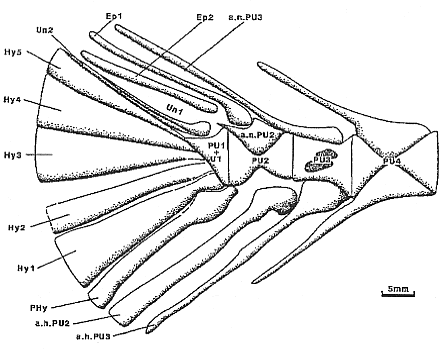 Figure 3 - Cliquez sur l'image pour accéder à la légende |
Le squelette caudal axial (fig. 3) est formé de trois éléments. Postérieurement, le complexe uro-terminal (PU 1 + U 1) est prolongé vers l'arrière par deux uroneuraux accolés redressés vers l'arrière qui déterminent avec l'axe de la colonne vertébrale un angle d'environ 140°. L'extrémité antérieure de l'uroneural antérieur (Un 1) est accolée dorsalement à la partie postérieure du complexe uro-terminal. Son extrémité distale effilée prend appui contre la région proximale du second uroneural (Un 2) qui surmonte le dernier hypural. Ventralement, le complexe uro-terminal supporte le parhypural (PHy), long et relativement étroit, dont la largeur maximale est comprise environ 7 fois dans la longueur. Au-dessus prend place l'hypural antérieur (Hy 1), triangulaire, relativement large puisque sa largeur maximale égale approximativement le tiers de sa longueur. Le second hypural (Hy 2) , de forme subrectangulaire, est relativement étroit, sa largeur maximale égalant approximativement la moitié de celle de l'hypural antérieur. La partie dorsale de la nageoire caudale est soutenue par trois hypuraux de taille décroissante, les deux plus. inférieurs (Hy 3, Hy 4) ayant une largeur maximale un peu inférieure à celle de l'hypural antérieur (Hy 1) , alors que l'hypural supérieur (Hy 5) est très étroit. Dorsalement, deux épuraux prennent place en avant et au-dessus des uroneuraux.
La partie proximale élargie de l'épurai antérieur (Ep 1) est visible juste au-dessus du centrum préural libre postérieur (PU 2). L'épural postérieur (Ep 2) est sensiblement plus court car son extrémité proximale se situe au-dessus du complexe uro-terminal. Le centrum préural antérieur (PU 3) supporte une neurapophyse et une hémapophyse très allongées qui supportent respectivement les rayons marginaux antérieurs situés dorsalement et ventralement à l'avant de la nageoire caudale.
De la nageoire dorsale, seule subsiste la partie postérieure. Elle débute par une épine relativement courte dont la longueur n'excédait probablement pas la moitié de celle des lépidotriches antérieurs de la nageoire. Le premier d'entre eux est seulement articulé alors que les 11 lépidotriches suivants sont à la fois articulés et bifurqués. On dénombre 12 ptérygophores au-dessous de la nageoire dorsale postérieure.
La nageoire anale est opposée à la moitié postérieure de la dorsale postérieure. Bien que leur état de conservation soit fort médiocre, on distingue à l'avant trois épines, la deuxième étant nettement plus robuste que les deux autres. En arrière prennent place 9 lépidotriches dont le premier n'est pas bifurqué. L'endosquelette de l'anale paraît avoir comporté 10 ptérygophores. Le plus antérieur, sensiblement plus long et plus robuste que les suivants, s'articule avec l'hémapophyse portée par le centrum postabdominal antérieur pour constituer un complexe hémaxanal.
De la ceinture scapulaire seul est observable un fragment de cleithrum. Rien ne subsiste de la nageoire pectorale.
Au contraire, les os pelviens et les nageoires pelviennes sont bien visibles, sans qu'il soit cependant possible d'en donner une description précise, exception faite de la présence d'une épine et de cinq rayons à la nageoire pelvienne.
Le corps est couvert d'écailles cténoïdes dont le champ antérieur porte généralement une dizaine de radii disposés en éventail. La ligne latérale occupe une position haute sur le flanc car elle est située à mi-distance entre la région dorsale des centra vertébraux et la base de l'épine située à l'avant de la nageoire dorsale postérieure. Elle s'abaisse ensuite progressivement vers l'arrière où elle est masquée par les centra vertébraux.
Affinités
De la description précédente, il ressort que les squelettes de poissons fossiles découverts dans les «grès de Moulas» peuvent être caractérisés comme suit :
- poissons de grande taille dont la longueur standard pouvait excéder 450 mm ;
- neurocrâne surmonté d'une crête sagittale longue et basse ;
- opercule prolongé postérieurement par une seule épine ;
- colonne vertébrale comportant environ 11 vertèbres abdominales et 14 vertèbres postabdominales ;
- nageoire dorsale dédoublée ; nageoire dorsale antérieure composée de 7 épines, dont la troisième est la plus longue (sa longueur égale approximativement 20 % de la longueur standard) ; nageoire dorsale postérieure comportant une épine et 11 lépidotriches ;
- nageoire anale formée de trois épines dont la seconde est à la fois la plus longue et la plus robuste, et de 9 lépidotriches ;
- nageoire caudale en forme de palette, composée de ?+I+8/7+I+? rayons.
Cette énumération conduit à rapprocher ces poissons fossiles de l'espèce actuelle Lares niloticus (Linné) dont Agassiz (1833) a publié une reconstitution du squelette dans le tome IV de ses Recherches sur les Poissons fossiles (fig. 4).
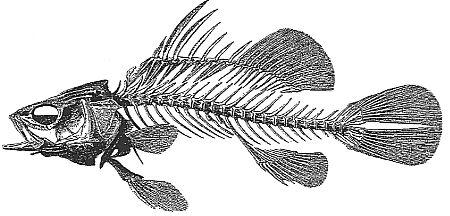 |
| Fig. 4 - Lates niloticus (Linné) - Reconstitution du squelette. D'après L. Agassiz (1833) |
Or, le genre Lates Cuvier & Valenciennes a été récemment subdivisé par Sorbini (1970) qui a attribué au genre Eolates Sorbini les espèces éocènes Lates gracilis Agassiz et L. macrurus Agassiz. C'est à ce même genre qu'il convient de rapporter également l'espèce L. aquensis Gaudant, de l'Oligocène supérieur d'Aix-en-Provence (Gaudant, 1981).
Bien que l'on connaisse relativement mal l'anatomie des poissons fossilisés dans les «grès de Moulas», il ne fait aucun doute qu'il s'agisse de représentants de la famille des Latidae, telle qu'elle est définie par Mooi & Gill (1995), cela notamment en raison de la possession d'une colonne vertébrale comportant plus de 24 vertèbres et d'un opercule prolongé postérieurement par une épine, deux critères utilisés par Greenwood (1976) pour distinguer Latinae (aujourd'hui Latidae) et Centropominae. Ces deux caractères ainsi que la morphologie de la crête sagittale du neurocrâne permettent en effet de distinguer les poissons des grès de Moulas, non seulement des représentants de la famille des Centropomidae, mais aussi du genre Cyclopoma Agassiz, un représentant de la famille des Percichthyidae apparenté au genre australien actuel Percalates Ramsay & Ogilby, auquel ils avaient été rapportés par Depéret (1912), très probablement au vu de leur grande taille. Or ces caractères conduisent à rapprocher les poissons de Moulas des genres Lates Cuvier & Valenciennes et Eolates Sorbini. Greenwood (1976) distinguait ces deux genres à l'aide des trois caractères suivants :
- squelette caudal axial d'Eolates Sorbini comportant, comme l'a montré Sorbini (1970, 1973), 3 épuraux (contre 2 chez Lates Cuvier & Valenciennes) et dont le second centrum préural porte une épine neurale bien développée (celle-ci est fortement réduite chez Lates) ;
- présence d'une seule ligne latérale sur la nageoire caudale chez Eolates Sorbini, alors que deux lignes latérales accessoires sont également présentes chez Lates Cuvier & Valenciennes ;
- nageoire dorsale moins profondément divisée chez Eolates Sorbini.
De ces trois caractères, un seul - l'anatomie du squelette caudal axial - a pu être observé de façon satisfaisante sur le matériel de Moulas. Or, la présence de seulement deux épuraux est décisive pour attribuer ces poissons au genre actuel Lates Cuvier & Valenciennes, cela bien qu'il n'existe aucune trace de lignes latérales accessoires sur la nageoire caudale et que la profondeur de l'échancrure entre les deux parties de la nageoire dorsale demeure inconnue sur les fossiles de Moulas.
 Figure 5 - Cliquez sur l'image pour l'agrandir |
Conséquences stratigraphiques
L'identification du genre Lates Cuvier & Valenciennes dans les «grès de Moulas» conduit à rajeunir considérablement l'âge de cette formation qui doit être rapportée au Miocène alors qu'elle était jusqu'alors attribuée à l'Eocène. En effet, alors que le genre Eolates Sorbini n'est actuellement connu en Europe que par deux espèces de l'Eocène moyen : E. gracilis (Agassiz), du Monte Bolca (Italie) et E. macrurus (Agassiz) du «Calcaire grossier» des environs de Paris (Sorbini, 1973), et par l'espèce E. aquensis (Gaudant) de l'Oligocène terminal d'Aix-en-Provence (Bouches-du-Rhône) (Gaudant, 1977), le plus ancien représentant européen du genre Lates Cuvier & Valenciennes décrit à ce jour est l'espèce L. partschii Heckel du Badenien (Miocène moyen) du bassin de Vienne (Autriche) (Heckel, 1856).
Signification paléoécologique
D'un point de vue paléoécologique, l'identification du genre Lates Cuvier & Valenciennes apporte peu d'informations relatives au milieu dans lequel vivaient les poissons fossilisés dans les «grès de Moulas». Il s'agit en effet soit de poissons euryhalins dont les espèces actuelles vivent principalement dans les eaux marines côtières et les estuaires, comme c'est le cas pour L. calcarifer (Bloch), soit de poissons dulcaquicoles largement répandus dans les fleuves et les grands lacs africains, que peuplent la perche nilotique, L. niloticus (L.), et six espèces endémiques des lacs Albert, Rodolphe et Tanganyika (Greenwood, 1976).
En revanche, la distribution géographique des espèces actuelles de ce genre (fig. 5) fournit une information significative sur le climat qui régnait dans les Pyrénées-Orientales lors du dépôt des «grès de Moulas». On constate en effet que, pour l'essentiel, leur aire de répartition se situe dans la zone intertropicale, ce qui correspond à des régions soumises à un climat à la fois chaud et humide.
La perche nilotique : un poisson vénéré par les anciens Egyptiens
Au sujet de la perche nilotique qui est connue à l'état fossile depuis la fin du Miocène (Otero & Sorbini, 1999), il est intéressant de rappeler que ce poisson fit l'objet, dans l'ancienne Egypte, d'un véritable culte de la part des habitants de la ville d'Esneh (en arabe Isna), en Haute-Egypte (entre Assiout et Assouan). Mais donnons plutôt la parole à Lortet, co-auteur de La faune momifiée de l'ancienne Egypte (1903) : «Les anciens Egyptiens semblent avoir eu la plus grande vénération pour un superbe poisson de la famille des Percoïdes, le Lates niloticus qui habite encore en quantités innombrables les eaux du Nil dans la Haute-Egypte. Certaines villes, entre autres Esné, vouaient un culte spécial à cette espèce ; non seulement les habitants honoraient le poisson vivant, mais encore par d'ingénieux procédés de momification, ils s'efforçaient de le préserver de toute destruction» (fig. 6). Pour cette raison, la ville d'Esneh reçut à l'époque de l'occupation gréco-romaine le nom de Latopolis.
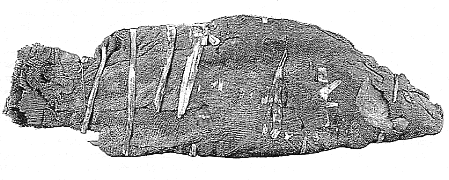 |
| Fig. 6. - lates niloticus (Linné). Momie provenant d'Esneh (Haute-Egypte), conservée au département des sciences de l'homme du Muséum d'histoire naturelle de Lyon sous le numéro d'inventaire 9000 1178. Longueur du poisson : 440 mm. |
Remerciements
L'auteur remercie M. P. Le Strat (BRGM, Montpellier) qui a suscité la présente étude. Il est également redevable envers le professeur R. Bourgat, conservateur du Muséum d'histoire naturelle de Perpignan, qui a autorisé le prêt des spécimens étudiés. Ses remerciements s'adressent également à Mme D. Emmons, conservateur du département des sciences de l'homme au Muséum d'histoire naturelle de Lyon, qui lui a communiqué le cliché d'une momie de lates conservée dans ce musée.
L'illustration a été préparée par M. J. Dyon.
Références
- AGASSIZ L., 1833-44. Recherches sur les poissons fossiles. Petitpierre, Neuchâtel, T. 4, VIII + 296 + 22 p.; atlas: pi. A-L + pl. 1-44.
- DEPERET C., 1912. Sur le grès éocène de Moulas, près Le Boulou (Pyrénées-Orientales). C. R. somm. Soc. géol. Fr., 1912 : 21-22, Paris.
- GAUDANT J., 1977. Additions à l'ichthyofaune stampienne d'Aix-en-Provence (Bouches-du-Rhône). Géol. méditerr., 4 : 205-210, Marseille.
- GAUDANT J., 1981. Mise au point sur l'ichthyofaune oligocène des anciennes plâtrières d'Aix-en-Provence (Bouches-du-Rhône). C. R. Acad. Sci., 292 (III) : 1109-1112, Paris.
- GAUDANT J., 1999. Présence du genre Lates Cuvier et Valenciennes (Poissons téléostéens, Percoidei) dans les grès de Moulas, près du Boulou (Pyrénées-Orientales). Géol. France, n° 4, 1999: 67-75.
- GREENWOOD P. H., 1976. A review of the family Centropomidae (Pisces, Perciformes). Bull. Brit. Mus. (Nat. Hist.), Zool., 29: 1-81, London.
- HECKEL J. J., 1856. Beitrâge zur Kenntniss der fossilen Fische Osterreichs. Denkschr. k. Akad. Wiss. Wien, Math.- natw. Cl., 11: 187-274, Wien.
- LORTET L. & GAILLARD C., 1903. La faune momifiée de l'ancienne Egypte. Arch. Mus. Hist. nat. Lyon, 8: I-VIII +1-205.
- Mool E. D. & GILL A. C., 1995. Association of epaxial musculature with dorsal fin pterygiophores in acanthomorph fishes and its phylogenetic significance. Bull. Nat. Hist. Mus., Zool., London, 61: 121-137.
- OTERO O. & SORBINI L., 1999. Etude systématique et anatomique du Lates (Perciformes, Latidae) du Messinien continental de Monte Castellaro, Italie. Considérations paléobiogéographiques. StudieRicerchesuiGiacimenti terziari di Bolca, Verona, 8:29-42.
- SORBINI L., 1970. Un nuovo genere fossile nell'ittiofauna di M. Bolca: Eolates nov. gen. Mem. Mus. civ. Storia nat. Verona, 18:11-29.
- SORBINI L., 1973. Evoluzione e distribuzione del genere fossile Eolates e suoi rapporti con il genere attuale Lates (Pisces : Centropomidae). Studi e Ricerche sui Giacimenti terziari di Bolca, Verona, 2:1-43.
Cet article a été publié dans les Annales du Muséum d'Histoire naturelle de Perpignan, n° 11, 2001 : pp.11-18